
Erythrina poeppigiana: shade tree gains new perspectives
FACT 96-06, September 1996
A quick guide to multipurpose trees from around the world
Erythrina poeppigiana (Walpers) O.F. Cook is a leguminous tree used in several agroforestry systems in Tropical America including shade for coffee, cacao and pastures, living fence posts, forage and fuel wood. It is also a promising species for alley cropping and mulching. Ease of management, high biomass production, nitrogen fixation and multiple uses make E. poeppigiana a suitable tree for farm and community forestry. It is known as “cámbulo” or “barbatusco” in Colombia, “bucare” or “cachimbo” in Venezuela, “amasisa” in Peru, “poró gigante”, “poró de sombra” or simply “poró” in Costa Rica, “pito” in Guatemala and Honduras, and “immortelle” or “mountain immortelle” in the West Indies; the more formal English name is “coral tree”. (Holdridge and Poveda 1975; Russo 1993).
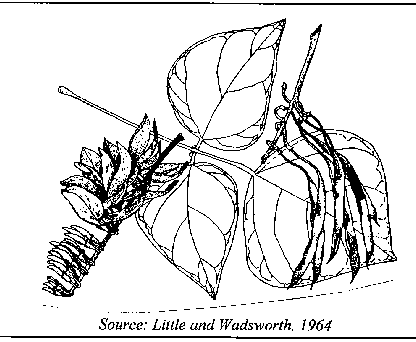 Botany
Botany
Erythrina poeppigiana belongs to the family Leguminosae, subfamily Papilionoideae, tribe Phaseoleae (Neill 1993). It is a large tree, growing to 35 m in height and 2 m in diameter. The crown is moderately spreading and the bole of large trees tends to be branchless below 10 – 20 m. The bark is grayish brown or gray, with thorn-like protuberances. Leaves are alternate, trifoliolate. The rhomboid-oval or oval folioles are 15 – 25 cm long and generally larger in saplings than in big trees. Glandular stiples below the paired lateral folioles are large and cup-shaped. Orange or reddish flowers are produced in racimes. The upper petal is wide and open. Erythrina poeppigiana is pollinated by perching passerine birds. The pods are 10-25 cm long. Seeds are brown, about 2 cm long and slightly curved. There are about 4,500 seeds per kg.
Ecology
Erythrina poeppigiana is native to humid and subhumid tropical lowlands, but cultivated and naturalized trees now are found to 2,000 m elevation (Holdridge and Poveda 1975). The average annual rainfall in its native and naturalized range is between 1,000 and 4,000 mm. In subhumid areas, it tolerates a 5-6 month dry season. Erythrina poeppigiana tolerates low soil fertility and relatively high acidity (down to pH 4.3), however tolerance varies by genotype (Pérez Castellón 1990).
In Costa Rica, the phenology of unpruned E. poeppigiana shifts from evergreen to deciduous along a rainfall gradient from the humid lowlands to the sub-humid mountains. The leafless period is quite short, and possibly caused by flowering rather than drought (Borchert 1980). A visible reduction of foliage during the flowering (December – January) also occurs under humid conditions. Pruning trees periodically will prevent complete leaf fall, and pruning trees once-a-year is enough to impede flowering (P. Nygren, pers. obs.).
Distribution
Erythrina poeppigiana is native to riverine and upland forests of the Amazon and Orinoco basins from Venezuela to Bolivia, and the moist Pacific forests of Ecuador and Colombia. It was introduced to Central America and a number of Caribbean Islands in the 19th century, and it has been widely naturalized in some areas like Costa Rica and Trinidad (Neill 1993).
Uses
Shade. Planted as a shade tree in cacao plantations in the humid tropics, E. poeppigiana conserves soil and contributes to high and sustainable cacao yields (Beer et al. 1990). Shade trees are partially pruned or not pruned at all. Production of N-rich litter (2.3 – 2.6%, Nygren 1995) is abundant, and the N supply in litterfall exceeds several times the export of N in the cacao harvest (Escalante et al. 1984).
In coffee plantations in Costa Rica, E. poeppigiana is usually pruned completely and lopped to a height of 2-3 m twice-a-year to promote coffee flowering and ripening of berries. The N supplied through pruning residues left on the ground fulfills the recommended N application rate for coffee in Costa Rica (Beer 1988). Farmers plant E. poeppigiana at spacings of 8 x 8 m and 6 x 6 m for unpruned and pruned trees, respectively.
Mulching and alley cropping. The green leaves of E. poeppigiana contain 4.1 – 4.9% nitrogen (Pérez Castellón 1990), which makes it an excellent species for green manure production. A ten-year experiment in Costa Rica measured the effects of cut-and-carry mulching with 20 tons/ha of E. poeppigiana fresh matter on maize and bean yields in a sequential cropping system. Crops were harvested once-a-year and production was good compared to local on-farm production. Crop production also increased each year of the experiment. The same experiment in Costa Rica evaluated alley cropping E. poeppigiana with maize and beans. Although satisfactory and sustainable for 10 years, the maize yield in this experiment was lower than the maize yield in the mulching experiment. The bean yield in the alley cropping system was both high and sustainable (Haggar et al. 1993). In a separate experiment in Costa Rica, E. poeppigiana alley cropping also sustained two maize crops per year over eight years without fertilization. Soil carbon and nitrogen pools decreased, but 50% less than in fertilized control plots (Dominique 1994).
For alley cropping, E. poeppigiana should be planted in dense hedgerows (1 – 2 m between trees), with wide alley (6 – 8 m) between tree rows (Kass et al. 1993a; Nygren and Jiménez 1993).
Forage. The green leaves of E. poeppigiana have a good nutritive value (20 – 22% of dry matter), are high in crude protein (27 – 34%) and have a good range of in vitro digestibility (49 – 57%). However, due to the high cell wall content (55 – 58%), they should be supplemented with energy sources, e.g. tropical grass, which are readily degradable in the rumen (Kass et al. 1993b). The presence of potentially toxic alkaloids in the leaves of E. poeppigiana has not affected the health of cattle or goats, but feeding leaves to non-ruminants may be risky (Kass 1994).
Other uses. The wood is light, with low calorific value bu it is sometimes used as fuel wood (Russo 1993).
Silviculture
Propagation. The seeds of E. poeppigiana may be stored for several years in tightly closed containers in a cool, dry place (ca. 5 °C, 30 – 40% relative humidity). Immersion in water at room temperature for 24 h enhances germination. The germination rate is about 70%. Germination takes 5-15 days. The seedlings may be planted in the field when they are 20-30 cm high (3 – 4 months), preferably at the beginning of rainy season. The seedling survival is generally good, but weed control may be necessary during the first year to enhance growth (Víquez and Camacho 1993; P. Nygren pers. obs.).
Air-layering to establish rooted cuttings yields a survival rate of 83% in vegetative propagation of E. poeppigiana. The roots appear about 6 weeks after air-layering. The leaves must be removed before planting, and the top cut made at a 45° angle and sealed with paraffin. Unrooted cuttings should be long (> 1.5 m). Stakes from lower and middle sections of one-and two-year-old branches give best results. Cuttings are planted at a depth of 30 cm. Inoculation of seeds or cuttings with Bradyrhizobium bacteria is not generally required in areas where E. poeppigiana is native or naturalized (Víquez and Camacho 1993; P. Nygren pers. obs.). However, inoculation is recommended when introducing the species to new areas.
Management. A formation pruning is recommended about 4-6 months after planting to remove the lowest branches. Normal pruning management may start 9-12 months after planting. Tall crops should not be associated with E. poeppigiana before the first complete pruning, but low crops may be planted at the time of the formation pruning. Coffee and cacao may be planted together with the trees. Due to the slow recovery of carbohydrate reserves, pruning of E. poeppigiana more often than twice-a-year causes the risk of debilitation and turnover of trees within a few years (Nygren et al. 1996).
Symbioses
E. poeppigiana nodulates abundantly with nitrogen fixing bacteria of genus Bradyrhizobium; peak values exceeding 1,000 kg/ha of nodules were reported for unpruned cacao shade trees, but during the driest season nodulation dropped to nil (Escalante et al. 1984). Globular nodules are formed in the site of lateral root emergence, and they have never been observed deeper than 10 cm. (Holdridge and Poveda 1975; Neill 1993; Víquez and Camacho 1993; P. Nygren, pers. obs.).
Soil acidity does not impede nodulation, but differences in the efficiency of bacterial strains were detected in a soil with 50% aluminum saturation (Gross et al. 1993). Pruning causes a complete turnover of nodules, and renodulation initiates about 2.5 months after pruning. After initiation, 66-180 kg/ha of nodules may be produced in a month (Nygren and Ramírez 1995).
Vesicular-arbuscular mycorrhizae improve nitrate uptake efficiency of unnodulated seedlings (Cuenca and Azcón 1994).
Limitations
Adult June beetles (Phyllophaga menetriesi, Coleoptera: Scarabaeidae) feed on young leaves of E. poeppigiana. Because June beetles lay eggs close to foraging areas, the root-eating larvae are a potential risk for associated crops (Hilje et al. 1993). Only minor damage to maize alley cropped with E. poeppigiana has been observed (D. Kass, pers. obs.), but the pest problem requires further investigation.
Selected references
Beer, J. 1988. Litter production and nutrient cycling in coffee (Coffea arabica) or cacao (Theobroma cacao) plantations with shade trees. Agroforestry Systems 7: 103-114.
Kass, D.C.L. 1994. Erythrina species – pantropical multipurpose tree legumes. In Gutteridge, R.C. and H.M. Shelton (eds). Forage tree legumes in tropical agriculture. CAB International, Wallingford, U.K. pp: 84 – 96.
Nygren, P. and C. Ramírez 1995. Production and turnover of N2 fixing nodules in relation to foliage development in periodically pruned Erythrina poeppigiana (Leguminosae) trees. Forest Ecology and Management 73: 59 – 73.
Westley, S.B. and M.H. Powell (eds) 1993. Erythrina in the New and Old Worlds. Nitrogen Fixing Tree Research Reports, Special Issue 1993. 358 p.
For a complete set of references contact the authors or FACT Net
Written by Pekka Nygren, INRA des Antilles-Guyane, Unite Agropedoclimatique, B. P. 515, 97165, Pointe-a-Pitre, Guadeloupe, French Antilles and Donald Kass, CATIE, 7170 Turrialba, Costa Rica
A publication of the Forest, Farm, & Community Tree Network (FACT Net)